Per cogliere le relazioni che il sistema motorio ha con i restanti sistemi (sensoriali, ma non solo) in cui si dispiega l'attività cerebrale è necessario spiegarne l'organizzazione.
A lungo si è ritenuto che i fenomeni sensoriali, percettivi e motori fossero ripartiti in aree corticali nettamente distinte: da un lato le aree sensoriali, articolate in aree visive (localizzate nel lobo occipitale), somatosensoriali (lobo parietale), uditive (circonvoluzione temporale superiore), ecc; dall'altro le aree motorie, situate nella parte posteriore del lobo frontale, nota anche come corteccia frontale agranulare.
Tra le prime e le seconde sono interposte vaste regioni corticali, definite come "aree associative": a esse (specie a quelle temporo-parietali), spetterebbe il compito di mettere insieme le informazioni provenienti dalle diverse aree sensoriali e di formare rappresentazioni oggettuali e spaziali da inviare alle aree motorie per l'organizzazione del movimento. Per esempio, se siete svegliati nel bel mezzo della notte da un forte rumore proveniente dall'esterno della camera da letto, dovete immediatamente decidere se agire o meno. Potreste cercare di determinare che cosa ha causato il rumore, se esso vi pone in una situazione di rischio e, in tal caso, che cosa fare per evitarlo. Potreste guardare all'esterno e fare attenzione a eventuali altri suoni insoliti. Per prendere una decisione, il nostro cervello deve elaborare informazioni provenienti dai sistemi sensoriali (dalla vista e dall'udito) e dalla memoria (cosa può generare un rumore come quello udito?). Molti di questi processi si verificano nelle aree associative.
Molte aree della corteccia cerebrale sono organizzate topograficamente in base alla loro funzione. Gli esempi più chiari di tale organizzazione topografico-funzionale sono rappresentati dalla corteccia motoria primaria (MI) situata nel lobo frontale e dalla corteccia somatosensoriale primaria (SI) posta nel lobo parietale. Le mappe dell'organizzazione di queste due aree corticali, in cui parti del corpo vicine sono rappresentate sulla superficie corticale in regioni vicine, sono definite homunculus motorio e homunculus sensitivo, scoperti da Wilder Penfield nella metà del Novecento.
Negli homunculus, ciascuna parte del corpo è mostrata sulla superficie della corteccia cerebrale che la rappresenta, e la dimensione relativa della parte corporea, è funzione delle dimensioni della relativa area corticale. Considerate le dita e il pollice: poiché essi sono capaci di movimenti precisi e fini e di un'elevata discriminazione tattile, le dita sono disegnate nella corteccia cerebrale con dimensioni superiori a quelle reali. Pertanto gli homunculus dimostrano che le aree della corteccia motoria, contenenti circuiti neuronali finalizzati ai movimenti dalle dita, e le aree della corteccia sensoriale contenenti circuiti neuronali relativi alle afferenze sensoriali provenienti dalle dita sono ampie. Altre aree della corteccia sono anch'esse topograficamente organizzate. Per esempio, si può trovare una mappa del campo visivo (cosa vedono gli occhi) nella corteccia visiva primaria o una mappa della frequenza dei suoni nella corteccia uditiva primaria1.
Il Sistema Motorio, risultava cosi suddiviso in tre parti:
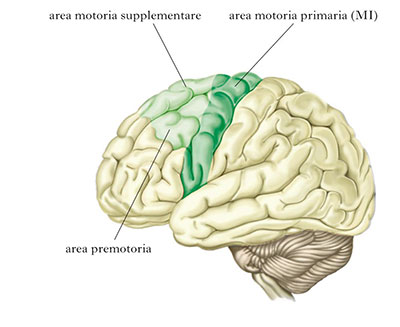
Ma ancor prima, agli inizi del Novecento, Korbinian Brodmann aveva costruito delle mappe citoarchitettoniche della corteccia della scimmia e dell'uomo.
Tali mappe furono ottenute da Brodmann grazie all'impiego di metodi istologici che consentivano di colorare selettivamente gli elementi cellulari e pertanto di distinguere le varie aree della corteccia cerebrale in base al numero degli stadi corticali (tipicamente sei), alla loro grandezza, alla quantità di neuroni contenuti e alla distribuzione dei tre tipi fondamentali di neuroni corticali (cellule piramidali, stellate, fusiformi).
Egli ripartiva questa regione del lobo frontale in due aree diverse: Area 4 e Area 6, sulla base della distribuzione delle cellule piramidali del V strato corticale.
Dalla sovrapposizione delle due scoperte si e visto come molte delle aree dell'homunculus non corrispondevano alle aree scoperte da Brodmann. Nonostante le critiche ricevute, lo schema generale dei due homunculus è stato, per molti anni, uno dei punti fermi della neurologia. E questo per almeno due ragioni: anzitutto, esso fornisce una spiegazione immediata e di facile applicazione al problema della localizzazione dei movimenti della corteccia motoria; inoltre, rispecchia l'idea, oggi come allora assai diffusa, dell'unitarietà funzionale del sistema motorio corticale, vero e proprio punto di arrivo dell'informazione sensoriale elaborata dalle aree associative e in sé privo di ogni valenza percettiva e cognitiva. Contrariamente a quanto sopra citato, la corteccia motoria risulta formata da una costellazione di regioni diverse2.
Se confrontiamo una moderna parcellizzazione anatomo-funzionale della corteccia agranulare, notiamo come la corteccia motoria primaria (MI o anche chiamata F1), coincide con l'area 4 di Brodmann. Per quel che concerne l'area 6, essa appare distinta in tre regioni diverse:
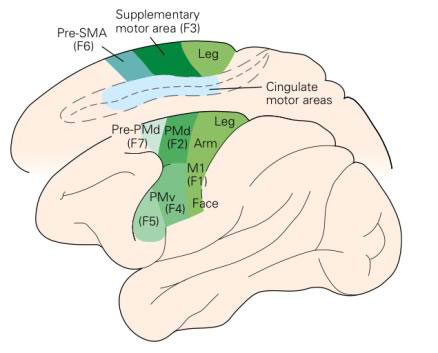
Ecco allora, come la scoperta che la corteccia agranulare possieda una struttura molto più complessa di quella ipotizzata in passato, consente di superare il distacco che vi è tra il sistema motorio da un lato e i sistemi sensoriali dall'altro. Per cogliere appieno natura e portata del sistema motorio corticale non basta però individuare i diversi tasselli che compongono il mosaico delle aree anatomicamente e funzionalmente distinte della corteccia agranulare. Occorre considerare anche le loro connessioni con le altre aree motorie (connessioni intrinseche), quelle con le aree corticali al di fuori della corteccia frontale agranulare (connessioni estrinseche), nonché l'organizzazione delle loro proiezioni ai centri sottocorticali e al midollo spinale (connessioni discendenti).
Per quel che concerne le connessioni estrinseche, le aree della corteccia frontale agranulare ricevono afferenze corticali da tre regioni principali:
Entrambi i lobi sono formati da una molteplicità di aree indipendenti, ciascuna delle quali appare deputata a elaborare determinati aspetti dell'informazione sensoriale e risulta connessa a fattori specifici. La corteccia motoria posteriore presenta, dunque, una parcellizzazione simile a quella riscontrata nella corteccia motoria.
Ma la cosa più importante è che nella aree parietali posteriori, a lungo classificate come aree associative, è stata osservata un'attività neurale in connessione ad atti motori. Per definizione, possiamo dire che sono motori quei neuroni la cui attività è collegata a un movimento; ciò vuol dire allora che la corteccia parietale posteriore deve essere considerata parte del sistema motorio corticale?
Non è proprio così: le aree parietali non sono altro che il punto di arrivo delle informazioni sensoriali provenienti da varie parti del corpo ma è attraverso i neuroni specchio che queste sensazioni riescono a essere trasportate alla corteccia frontale per essere tradotte in movimenti. Quindi potremmo considerare questi neuroni come "un ponte" attraverso cui l'informazione sensitiva passa dal lobo parietale a quello frontale e diviene movimento.
Sul piano funzionale ciò si traduce nel fatto che ciascuno di questi circuiti appare coinvolto in una particolare "trasformazione" sensori-motoria, ossia in una particolare "traduzione" di una discriminazione dello stimolo sensoriale in termini motori.
Le aree motorie posteriori, infatti, ricevono le loro principali afferenze corticali dal lobo parietale. Le aree motorie anteriori, invece, dalla corteccia prefrontale e da quella del cingolo. Le aree posteriori ricevono dal lobo parietale una ricca messe di informazioni sensoriali che utilizzano per l'organizzazione e il controllo del movimento, elaborandole attraverso una serie di processi paralleli che vedono ogni circuito coinvolto nell'effettuazione di specifiche trasformazioni sensori-motorie.
Alle aree anteriori, invece, giungono scarse informazioni sensoriali, ma tali aree recepiscono informazioni cognitive di ordine superiore, legate alla pianificazione temporale delle azioni o alle motivazioni. Esse hanno principalmente funzioni di controllo, determinando in particolare quando e in quali circostanze il movimento selezionato dalle aree posteriori debba tradursi in un atto effettivo3.