Innanzitutto è stato esaminato se, quando l'esercizio era combinato con il digiuno, ci fosse un maggior aumento delle proteine correlate all'autofagia (Fig1) e dei geni correlati all'autofagia (Fig.2). Come marker della presenza di AP nella cellula è stata considerata la forma lipidated di LC3b (LC3b-II), già ampiamente usata (Barth S, et al., 2010), mentre per valutare la sintesi di AP è stato determinato di rapporto di LC3b-II sulla forma non-lipidated(LC3b-I), considerato come un test affidabile (Klionsky DJ, 2007). LC3b-II era più alto dopo l'esercizio o il digiuno e la combinazione dei due stimoli ha determinato un ulteriore aumento di LC3b-II (Fig.1B). La forma non-lipidated di LC3b(LC3b-I) era drasticamente ridotta con il digiuno e non era influenzata successivamente dall'esercizio (Fig.1A). Come conseguenza, il rapporto LC3b-II/I era drammaticamente aumentato in risposta al digiuno (Fig.1C). Lc3bm RNA era aumentato a seguito dell'esercizio nella condizione di digiuno, ma non in quella di sazietà (Fig.2A). Benchè Gabarapl1 mRNA è risultato elevato in vari modelli di atrofia associata ad un aumento dell'autofagia (Mammucari C et al., 2007; Zhao J, et al., 2007), la determinazione dell'espressione della proteina è meno comune. Come per la sottofamiglia LC3, l'attaccamento dei membri della sottofamiglia Gabarap alla membrana AP dipende dalla loro coniugazione alla fosfodietiletanolamina (Kabeya Y, et al., 2004). Contrariamente a LC3b, l'aumento del rapporto Gabarapl1-II/I nel gruppo Digiuno+Corsa (Fig.1F) era dovuto solamente alla deplezione della forma non-lipidated (Fig.1D), mentre la forma lipidated era rimasta inalterata (Fig.1E). Similarmente a Lc3b, Gabarapl1 mRNA era più alto quando digiuno ed esercizio erano cumulati.
I complessi Atg12-Atg5-Atg16L erano specificamente legati al fagoforo (doppia membrana che precede la formazione dell'autofagosoma) ma dissociati dopo il completamento della formazione dell'AP (Mizushima N, et al., 2003a; Mizushima N, et al., 2003b). Quindi, un aumento nella presenza di questi complessi può essere usato come marker del fagoforo ( Weidberg H, et al., 2011). Nel presente studio, Atg5 forma coniugata di Atg12 (cAtg12) era aumentata solo quando l'esercizio era combinato con il digiuno (Fig.1G). L'Atg4b proteasi ha un'ampia specificità verso tutti gli Atg8 di mammifero(Hemelaar J, et al., 2003) ma preferenzialmente scinde LC3 (Kabeya Y, et al., 2004). Atg4b in questo esperimento tendeva a diminuire a seguito del digiuno (Fig.1H; P=0.065), possibilmente a causa di una ridotta delipidizzazione di LC3b.
L'accumulo di LC3b-II all'interno della cellula può essere dovuto a difetti nella degradazione da lisosoma, che può essere valutata attraverso l'accumulo di p62/SQSTM1 (Mizushima N, and Yoshimori T, 2007). In questo esperimento, p62/SQSTM1 è rimasto stabile nei gruppi Sazietà+Corsa e Digiuno+Riposo e tendeva a diminuire nel gruppo Digiuno+Corsa (P=0.079, Fig.1I). Questo trend è stato accoppiato ad un aumento della p62 mRNA nel gruppo Digiuno+Corsa (Fig.2C), e questo risultato fa supporre che vi è stato un aumento nel turnover di p62/SQSTM1.
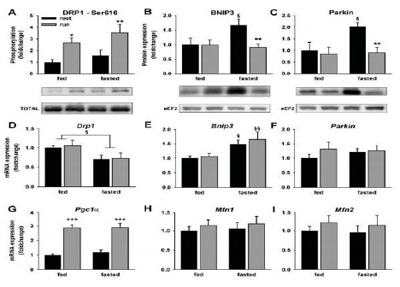
Gli sperimentatori hanno poi valutato se l'aumento osservato dei markers dell'autofagia potesse essere associato a variazioni del rimodellamento dei mitocondri e dei markers della mitofagia. Si è osservato che l'esercizio ha indotto la fosforilazione di DP1Ser616 promuovendo, potenzialmente, la fissazione mitocondriale attraverso la translocazione di DRP1 alla membrana mitocondriale (Taguchi N, et al., 2007) (Fig.3A). DRP1 mRNA non era modificato dall'esercizio, ma era diminuito nella condizione di digiuno (Fig.3D). E' noto che BNIP3 promuove la mitofagia interrompendo il potenziale di membrana mitocondriale (Sandoval H, et al, 2008). Nell'esperimento in questione, la proteina BNIP3 era aumentata a seguito del digiuno negli animali a riposo ma questo aumento è stato completamente invertito dall'esercizio (Fig.3B). Poiché Bnip3 mRNA era anche più alto nella condizione di digiuno ed è rimasto elevato dopo l'esercizio (Fig.3E), è stato ipotizzato che il digiuno abbia aumentato l'espressione di BNIP3, che veniva invece degradata durante l'esercizio attraverso una più alta attività mitofagica. Similmente, Parkin, che è una ubiquitinligasi che media la ubiquilazione dei mitocondri (Youle RJ and van der Bliek AM, 2012) era più alta nel gruppo Digiuno+Riposo mentre l'esercizio ha causato una rimozione di questa proteina (Fig.3C). Parkin mRNA è rimasta inalterata in tutte le condizioni (Fig. 3F). Il principale regolatore della biogenesi mitocondriale Pgc1a era più alto dopo l'esercizio, ma non era alterato dallo stato nutrizionale (Fig.3C). I markers di fusione mitocondriale Mfn1 e Mfn2 non sono risultati influenzati né dal protocollo di esercizio né dallo stato nutrizionale degli animali (Fig.3H, I).
Figura 1. Cambiamenti nelle proteine legate all' autofagia in risposta ad esercizi svolti su tapis roulant eseguiti sia in condizioni di digiuno, sia in condizioni di sazietà. I valori sono presentati come medie ± ES. *p< 0.05 , **p< 0.01 , ***p< 0.001 confrontati a riposo (effetto dell' esercizio). §p< 0.05 , §§p< 0.01 , §§§p< 0.001 confrontati con lo stato di sazietà (effetto del digiuno). Wester blots rappresentativi sono aggiunte sotto ogni variabile. Aggiustamento per contrasto e brillantezza era applicato all'intera figura.
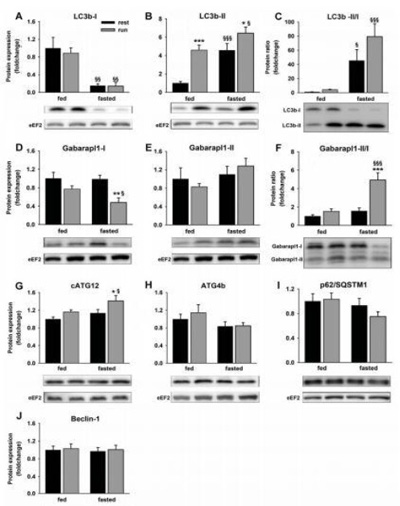
Figura 2. Cambiamenti nei geni correlati all' autofagia in risposta ad esercizi svolti sul tapis roulant eseguiti sia in condizioni di digiuno, sia in condizioni di sazietà. I valori sono presentati come medie ± ES. *p< 0.05 , confrontati a riposo (effetto dell' esercizio). §p< 0.05 , §§p< 0.01 , §§§p< 0.001 confrontati con lo stato di sazietà (effetto del digiuno).

Figura 3. Cambiamenti nella fissione, mitofagia, biogenesi mitocondriale e nella fusione dei markers correlati all' autofagia in risposta ad esercizi svolti sul tapis roulant eseguiti sia in condizioni di digiuno, sia in condizioni di sazietà. I valori sono presentati come medie ± ES. *p< 0.05 , confrontati a riposo (effetto dell' esercizio). §p< 0.05 , §§p< 0.01 , confrontati con lo stato sazietà (effetti del digiuno). Appropriate Wester blots rappresentativi sono aggiunte sotto ogni variabile. Aggiustamento per contrasto e brillantezza era applicato all'intera figura.
Il proposito secondario del presente studio era spiegare i meccanismi molecolari sottostanti la più alta attivazione dell'autofagia quando l'esercizio veniva effettuato in una condizione di digiuno. L'autofagia è trascrizionalmente attivata da FoxO3a e inibita post-translazionalmente dal target mammifero di rapamicina (mTORC1) (Kim J, et al., 2011; Mammucari C, et al., 2007). Entrambe le proteine sono controllate dalla via insulina/fosfoinositide-3-kinasi(PI3K)/Akt. Nell'esperimento, l'esercizio ha fortemente ridotto la concentrazione plasmatica di insulina (Fig.4A), la quale non era rilevabile quando l'esercizio e il digiuno erano combinati, e quindi la sua concentrazione era più bassa di 4.35 pM, come specificato dal produttore (Mercodia). Similmente al pattern dell'insulina, lo stato di fosforilazione di Akt sia su Ser473 (Fig.4B) che Thr308 (Fig.4C) era più basso nel gruppo Digiuno + Corsa. Akt previene la traslocazione nucleare di FoxO3a fosforilando i residui multipli di questo fattore di trascrizione tra i quali vi è Thr32 (Greer El and Brunet A, 2005). Nel gruppo Digiuno + Riposo è stato osservato un aumento in FoxO3aThr32 (Fig.4E). Conseguentemente, il rapporto tra FoxO3aThr32 e la sua forma totale era diminuito quando l'esercizio era condotto in una condizione di digiuno (P=0.004, dati non mostrati), supportando l'idea di una traslocazione nucleare di FoxO3a in questa condizione. Per valutare l'attività di mTORC1 gli autori hanno misurato lo stato di fosforilazione di S6K1 e 4E-BP1, due target a valle di mTORC1. S6K1Thr389 era drammaticamente diminuito a seguito dell'esercizio e del digiuno (Fig.4F) e lo stato di fosforilazione di 4E-BP1Thr37/46 era più basso nel gruppo Digiuno + Corsa (Fig.4G), suggerendo che l'attività di mTORC1 era più ridotta in questi animali. mTORC1 è anche una chinasi per ULK1Ser757, e la sua attività regola l'induzione dell'autofagia (Kim J, et al., 2011). ULK1Ser757 era ampiamente de-fosforilata in risposta all'esercizio combinato con il digiuno (Fig.4H). La forma totale di ULK1 era up-regolata dall'esercizio e dal digiuno ma i due stimoli non era addizionali (Fig.4I). L'autofagia può anche essere attivata dal sensore di energia AMPK, che tra l'altro fosforila ULK1 in più siti (Greer EL et al., 2007; Kim J, et al., 2011). Nell'esperimento in questione, lo stato di fosforilazione di AMPKaThr172 non era cambiato (Fig.4J), così come non erano ambiati i due siti di fosforilazione AMPK-dipendenti di ULK1, precisamente ULK1Ser555 (Fig.4K) e ULK1Ser317 (Fig.4L).
Figura 4. Cambiamenti nei componenti della trasduzione del segnale in risposta a ad esercizi svolti sul tapis roulant eseguiti sia in condizioni di digiuno, sia in condizioni di sazietà. I valori sono presentati come medie ± ES. *p< 0.05 , confrontati a riposo (effetto dell' esercizio). §p< 0.05 , §§p< 0.01 , confrontati con lo stato di sazietà (effetti del digiuno). Appropriate Wester blots rappresentativi sono aggiunte sotto ogni variabile. Aggiustamento per contrasto e brillantezza era applicato all'intera figura.

Altri segnali di trasduzione dello stress cellulare potrebbero potenzialmente essere responsabili dell'osservato aumento dell'autofagia. p38 MAPK è attivato da una varietà di stress tra i quali l'esercizio. E' stato dimostrato che p38 MAPK lega lo stress ossidativo all'attivazione dell'autofagia (McClung JM, et al., 2010). La fosforilazione di p38Thr180/Tyr182 era più alta a seguito dell'esercizio nella condizione di sazietà ma non era aumentata ulteriormente con l'esercizio nello stato di digiuno (Fig.5A). I carbonili proteici invece sono rimasti inalterati (Fig.5B).
Figura 5. (A) lo stato fosforilato di P38 Thr180/Tyr182 ; (B) livelli del carbonile proteico in risposta ad esercizi svolti sul tapis roulant eseguiti sia in condizioni di digiuno, sia in condizioni di sazietà. I valori sono presentati come medie ± ES. ***p< 0.001 confrontati a riposo (effetto dell' esercizio). §§p< 0.01 confrontati con lo stato di sazietà (effetto del digiuno). Appropriate Wester blots rappresentativi sono aggiunte sotto ogni variabile. Aggiustamento per contrasto e brillantezza era applicato all'intera figura.

Lo stress del reticolo endoplasmatico (ER) è attivato nel muscolo scheletrico in risposta all'esercizio di endurance (Jamart C, et al., 2012; Wu J, at al, 2011). Per far fronte allo stress dell'ER, la cellula attiva una risposta non incrociata della proteina (UPR).
Gli effettori a valle di questa via - tra i quali eIF2a – inducono l'espressione dei fattori di trascrizione come l'attivazione del fattore 4 (ATF4) di trascrizione e la proteina omologa (CHOP) della proteina di legame CCAAT/enhancer (C/EBP), così come l'unione ad incastro della proteina1 legante il fattore X di trascrizione (XBP1s). E' noto che ATF4 gioca un ruolo nella trascrizione di LC3 a Atg2 creando il legame tra ER-stress e autofagia (Kouroku Y, et al., 2007; Rouschop KM, et al., 2010). Nell'esperimento, l'esercizio ha aumentato il livello di Atf4 mRNA ma i test post-hoc all'interno di ogni condizione nutrizionale non sono risultati significativi (Fig.6A). La fosforilazione di eIF2aSer51 era aumenta quando l'esercizio veniva effettuato in una condizione di sazietà, ma non nella condizione di digiuno (Fig.6B). L'esercizio ha aumentato Xbp1s mRNA nel topo sazio ma non significativamente nel topo a digiuno (Fig.6C). Chop mRNA era più basso nella risposta all'esercizio in condizione di sazietà, mentre il digiuno ha ridotto Chop sia nel gruppo a riposo che in quello running (Fig. 6D).
Figura 6. Cambiamenti nei markers del ER-stress in risposta ad esercizi svolti sul tapis roulant eseguiti sia in condizioni di digiuno, sia in condizioni di sazietà. I valori sono presentati come medie ± ES.*p< 0.05 ***p< 0.001 confrontate a riposo (effetto dell' esercizio). §p< 0.05 , §§p< 0.01 , §§§p< 0.001 confrontate con lo stato di sazietà (effetto del digiuno). Appropriate Wester blots rappresentativi sono aggiunte sotto ogni variabile. Aggiustamento per contrasto e brillantezza era applicato all'intera figura.
